
Een crossing-over, ook wel recombinatie genoemd bij zebravinken, is een herindeling van de genetische eigenschappen. Een crossing-over komt regelmatig voor bij zebravinken, echter het gewenste resultaat (de juiste combinatie van genen) kan soms jaren op zich laten wachten.
Om iets meer van de erfelijkheid te begrijpen, zal ik eerst wat meer uitleggen over de vererving van een zebravink. Een zebravink heeft 9 chromosoomparen (tweetallen), terwijl een mens er 23 heeft. Een zebravink heeft dus in totaal 18 chromosomen. Een chromosoom bestaat weer uit twee helften, genaamd chromatiden, die om elkaar heen gedraaid zitten.
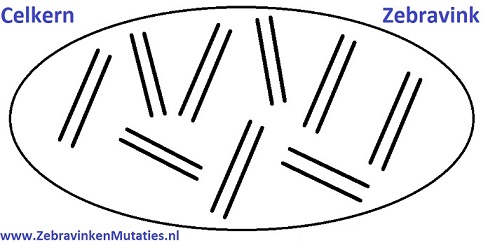
Een chromosoom bevat veel genetische informatie, zo zouden er meerdere mutaties aanwezig kunnen zijn op hetzelfde chromosoom. Zo zijn eigenschappen zoals kleur, model en formaat ook vastgelegd in de genen. Een (kleur)mutatie van een zebravink bevindt zich op een chromosoom. Deze mutatie ligt ergens vastgelegd in een gen op de chromatiden van de chromosoom, een soort lange slinger aan genetische informatie (genen).
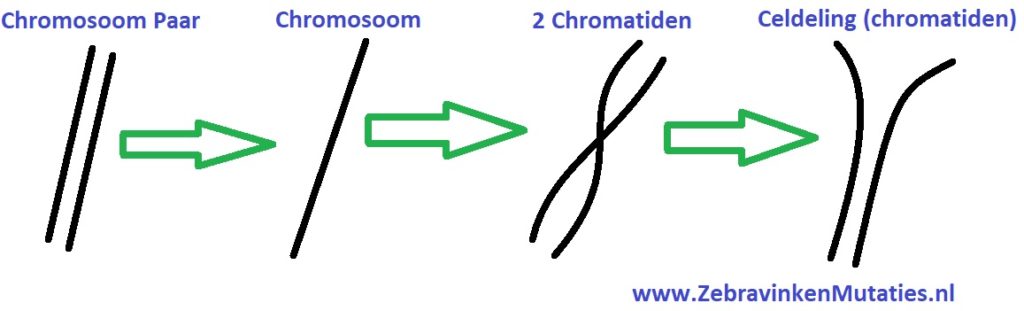
Bij een celdeling van de zebravink delen de 18 chromosomen zich in (36) chromatiden, vervolgens splitsen de chromatiden zich in 2 groepen. Na de splitsing reproduceren de chromatiden zich en ontstaan er nieuwe chromosomen (paren), nu zijn er dus 2 cellen welke een kopie van elkaar zijn.
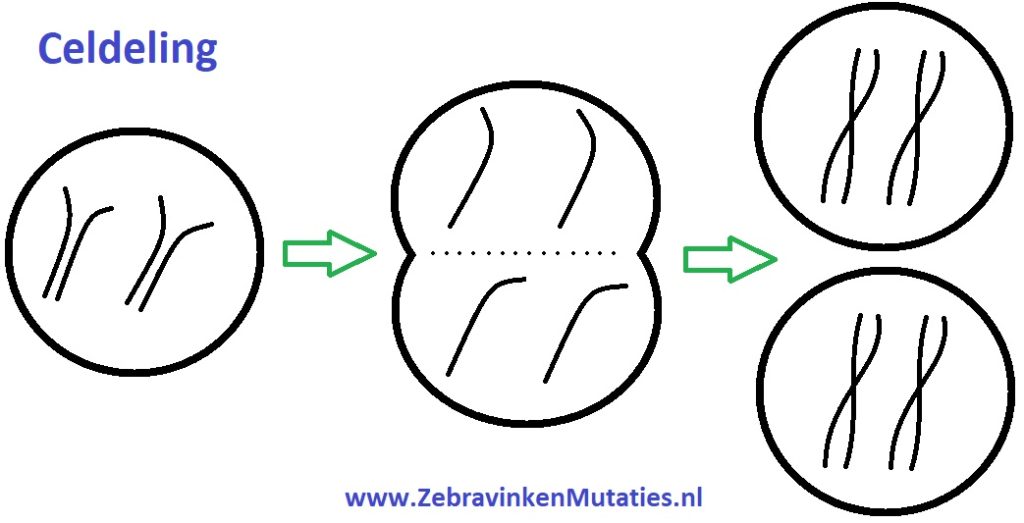
Een Crossing-Over (recombinatie) ontstaat echter al vóór de celdeling.
Bij de bevruchting komt een mannelijke en een vrouwelijke gameet (zaadcel en eicel) samen en smelten deze samen tot één cel. Een gameet (geslachtscel) bezit alleen enkelvoudige chromosomen (en dus geen paren). De gameet van het mannetje smelt dus samen met de gameet van het vrouwtje en vormt op deze manier weer chromosoom paren.
Vóór de celdeling is er veel beweging in de celkern waardoor de chromosomen (van de man en pop) regelmatig afbreken. Deze chromosomen zoeken elkaar echter weer op en groeien in bijna alle gevallen weer aan elkaar, ze worden tot elkaar aangetrokken. Echter heel soms komt het voor dat de 2 chromosomen op exact dezelfde locatie afbreken en aan het andere chromosoom van hetzelfde paar vastgroeien (dus die van de man, met die van de pop). Zodoende kan een mutatie van het ene chromosoom op het andere chromosoom terecht komen.
Vervolgens begint de celdeling, omdat de éérste cel gemuteerd is en alle volgende cellen kopieën zijn van deze eerste cel, zal elke toekomstige cel van de zebravink deze eigenschap bij zich dragen.
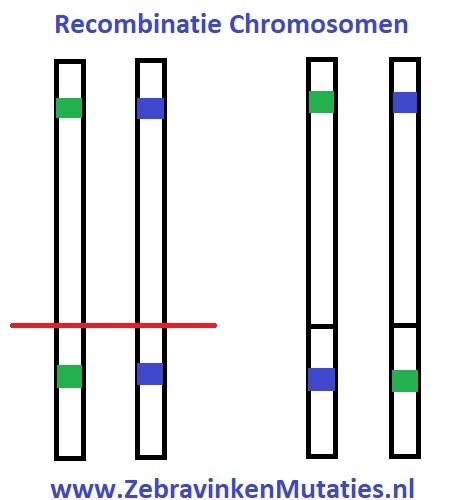
Er komt in werkelijkheid nog veel meer bij kijken bij een crossing-over. Echter als ik té ver in detail ga, zal het voor de meeste mensen niet meer begrijpelijk zijn, dus ik probeer het bij een zo simpel mogelijke basis te houden. Een crossing-over komt regelmatig voor, maar hier merken we eigenlijk niks van. Er veranderd qua uiterlijk in de meeste gevallen niks aan de zebravink. Pas wanneer 2 gemuteerde factoren (genen) op één chromosoom terecht komen kan het interessant worden.
Voorbeeld Crossing-Over Bleekrug Bruin
De mutatie Bleekrug en Bruin lagen beide in het geslachtschromosomen paar, echter niet op hetzelfde chromosoom. Hierdoor kon een zebravink qua uiterlijk nooit bleekrug en bruin tegelijk zijn. Echter door een recombinatie kwamen deze 2 genen op hetzelfde chromosoom terecht en ontstond de Bleekrug Bruine zebravink. Deze crossing-over was doelgericht geprobeerd te behalen, toen het na enkele jaren nog niet gelukt was werden alle vogels bij elkaar in een volière gegooid. Bij deze groep vogels is toen spontaan één Bleekrug Bruine man geboren.
Deze jonge bleekrug bruine man was afkomstig van een grijze split Bleekrug en Bruine man met een Masker Bruine pop. Er heeft dus een crossing-over plaatsgevonden tussen de gameet (geslachtscel) van de man, met de gameet van de pop. De man kon enkel bleekrug óf bruin doorgeven, omdat deze niet op hetzelfde chromosoom lagen. In dit geval moet deze man dus bleekrug doorgegeven hebben in het geslachtschromosoom, de pop gaf maskerbruin door.
Bij de crossing-over zijn beide chromosomen van man en pop gebroken, tussen het masker en bruin van de pop in. Vervolgens groeide de 2 chromosomen weer aan elkaar, echter aan het andere chromosoom van hetzelfde paar. Hierdoor kwam bruin en bleekrug op hetzelfde chromosoom te liggen (en masker zou in theorie nu weer alleen op het andere chromosoom liggen, deze was dus ontkoppeld).
Echter deze crossing-over (zoals sommige artikelen en boeken beschrijven) had in theorie niet direct zichtbaar geweest kunnen zijn, maar aanwezig als split BleekrugBruin en split masker (grijs). Omdat het jong een man was en deze dus BleekrugBruin op het ene chromosoom en Masker op het andere chromoom had. (en qua uiterlijk dus grijs had moeten zijn)
De Crossing-over was al niet zichtbaar aanwezig!
Echter bleef het niet bij één, maar werden er méérdere bleekrug bruinen mannen geboren uit dit koppel. Ik heb me helemaal stuk gelezen en alle scenario’s uitgedacht, maar ik kan maar tot één conclusie komen. Het zou daarnaast in theorie vrijwel onmogelijk zijn als dezelfde crossing-over meerdere keren bij één nestje voorkomt.
De vader van de Bleekrugbruine mannen, een grijze man welke geacht werd split bleekrug en split bruin te zijn, was dus al split BleekrugBruin. Deze grijze man was de stamvader van alle bleekrug bruine vogels, de crossing-over had bij deze man al plaatsgevonden en was als split kenmerk aanwezig, NIET zichtbaar!
De ouders van deze grijze man waren een bruine en een bleekrug grijze vogel. Zoals hierboven al uitgelegd, zaten de bleekrug en bruin dus op een ander chromosoom, echter vóór de celdeling vond de crossing-over plaats, waarbij bleekrug en bruin op hetzelfde chromosoom kwamen te liggen. Echter doordat het maar op één chromosoom lag, was dit alléén aanwezig als splitkenmerk. Doordat deze vervolgens gepaard werd aan een maskerbruine, kwamen hier dus Bleekrug bruine split Masker bruine mannen uit. Dit omdat bleekrugbruin en maskerbruin samen in de MM-reeks liggen, en bleekrug dominant is over masker. (in theorie hadden er dus ook direct volle bleekrugbruine poppen uit deze grijze man kunnen komen)
Kans op Crossing-Over
De kans op een gewenste Crossing-Over kan enorm variëren. Dit is afhankelijk van de afstand tussen de gemuteerde genen op het chromosoom. Stel dat er 30 genen tussen de 2 gemuteerde genen zitten, is de kans 30x groter dan wanneer de gemuteerde genen direct naast elkaar liggen. Zodoende kan het soms vele jaren duren voordat een gewenste recombinatie behaald wordt.
Meerdere soorten crossing-overs
Overigens is er niet maar één soort crossing-over, in werkelijkheid zijn er 4 vormen van crossing-overs. (Single-Crossover, Double-Crossover, Three-strand Double Crossover (two ways) en Four-strand Double crossover.) Waarbij de chromatiden dus bij een of meerdere delen afbreken en verkeerd aan elkaar groeien, waardoor compleet gemixte nieuwe chromosomen kunnen ontstaan. Een crossing-over kan dus invloed hebben op meestal 2, soms 3 en soms zelfs alle 4 de chromatiden, van het ouderpaar.
Hierboven vertelde we dat de afstand tussen de genen op het chromosoom ook invloed heeft, bij de kans op een crossing-over. Hierbij is het goed om te weten dat bij crossing-overs, in MAXIMAAL 50% van de gevallen een recombinatie kan optreden. Het maximaal haalbare is dus wanneer de genen zo ver mogelijk uit elkaar geplaatst zijn op het chromosoom. Oftewel, bij minimaal 50% van de crossing-overs die plaats vinden, merken we daar (qua uiterlijk) niks van. Deze crossing-overs (zonder invloed op mutaties) zorgen er mede voor dat elk individu uniek blijft, dit is goed voor de genenpoel.
Eumo en Bevederingsfout
In theorie is het dus ook mogelijk om gekoppelde factoren in een chromosoom van elkaar los te koppelen. De eumo en de fout in de bevedering zijn in theorie 2 verschillende gemuteerde cellen op hetzelfde chromosoom. Echter moet er dus een keer een crossing-over plaatsvinden waarbij de chromosomen op exact de juiste locatie doorbreken (tussen de 2 gemuteerde cellen in) en aan het andere chromosoom vastgroeien.
Indien deze 2 gemuteerde cellen een redelijke afstand van elkaar op hetzelfde chromosoom hebben, dan is de kans dus groot dat er binnenkort ergens een Eumo zebravink wordt geboren zonder Bevederingsprobleem, of andersom. Maar stel dat de Eumo en de bevederingsfout vlakbij of direct naast elkaar op het chromosoom liggen is de kans op een crossing-over van deze 2 mutaties zéér klein. Maar vroeg of laat zal het toch een keer kunnen gebeuren?
Echter volgens mijn eigen theorie welke hierboven bij de BleekrugBruin geschreven is, ligt het in dit geval een stuk gecompliceerder. We hebben hier namelijk te maken met een autosomaal recessieve vererving, en géén geslachtsgebonden vererving en MM-reeks. Een crossing-over werkt anders bij een geslachtsgebonden vererving.
Crossing-over en Geslachtsgebonden
Er is nog één zeer belangrijk detail bij een geslachtsgebonden vererving. Bij een geslachtsgebonden vererving vindt nog een extra stap plaatst in het celproces, de meiose fase. In deze fase wordt ook een stofje aangemaakt die ervoor zorgt dat de chromosomen veelvoudig afbreken en vervolgens weer aan elkaar groeien. De kans op een crossing-over is daarom 1.000 x groter bij een geslachtsgebonden vererving ten opzichte van een autosomaal recessieve ververing, waarbij er geen meiose plaats vind, maar enkel mitose. Bij mitose is de kans op een crossing-over dus 1.000 x kleiner dan bij meiose.
Naast bovenstaand feit, kan er ook géén ontkoppeling plaatsvinden bij het kweken van eumo x eumo. Aangezien beide chromosomen gelijk zijn, al breken ze af en groeien ze vast aan het andere chromosoom, dan houden ze dezelfde eigenschap. Ook bij alle volle eumo’s welke direct uit split x split of vol x split geboren worden, kan nooit de gewenste crossing-over plaatsvinden. Je kan dus NOOIT direct een volle eumo kweken zonder veerdefect, hiervoor dient de eigenschap eerst als splitkenmerk aanwezig te zijn!
Alléén bij een splitvogel kan een crossing-over hebben plaatsgevonden waarbij het “veerdefect” en de “eumo” gen van elkaar losgekoppeld zijn en op het andere chromosoom terecht gekomen zijn. Deze vogel zal dus split zijn voor eumo, en daarnaast ook split voor het veerdefect. Helaas is dit alleen niet zichtbaar voor het menselijk oog!
Indien er dus ooit een jong met énkel het veerdefect geboren wordt, dient er JUIST verder gekweekt te worden met deze ouders, want één van de ouders (óf mogelijk zelfs de voor-ouders!) is een splitvogel welke zowel eumo als het veerdefect onafhankelijk van elkaar kan vererven! Uit ditzelfde koppel kan dus een jong geboren worden welke een volle eumo is, split voor het voordefect! Door gericht te kweken en te selecteren kan je vervolgens een stam eumo opbouwen, zonder het veerprobleem.
Hoe kan je deze crossing-over bereiken?
Dit kan dus alleen bij het kweken van splitvogels, de kans op splitvogels is gelijk bij alle volgende combinaties: Split x split (50% = split), split x vol (50% = split) en split x normaal (50% = split). Het lastige is dat je nooit zult weten welke splitvogel de mogelijke gewenste crossing-over bevat, tenzij je ermee gaat kweken.
Mogelijk bestaan er dus al zebravinken welke split voor eumo en het veerprobleem (onafhankelijk van elkaar zijn) maar weten wij dit niet. Het is zelfs mogelijk dat deze crossing-over al meerdere keren is ontstaan, maar deze splitvogels niet gebruikt zijn voor de kweek, waardoor we het nooit zullen weten…
© Jesse van Es
www.ZebravinkenMutaties.nl
Gepubliceerd 10-2-2019